|
33. Lycaena dispar (Haworth, 1803) / Large copper / Lycaenidae – Lycaeninae
NL: grote vuurvlinder / D: Groβer Feuerfalter / F: cuivré des marais, le grand cuivré
    Photographs: Frits Bink ©. Photographs: Frits Bink ©.
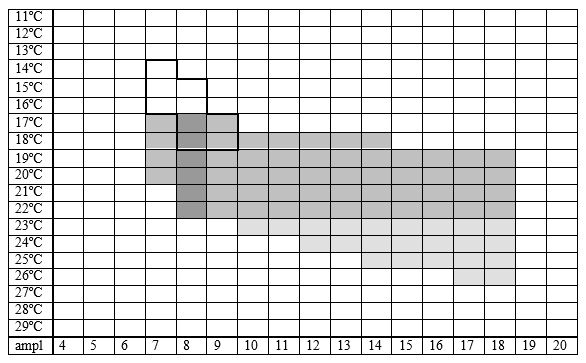
Lycaena dispar batava (Oberthür 1923), medium-sized, wing length 21 (19-23) mm; L. dispar carueli (Le Moult 1945), small, wing length 17 (16-18) mm; L. dispar rutila (Werneburg 1864) small or medium-sized, wing length 19 (17-22) mm. Butterflies of second brood are always smaller than of the first one. The species occurs in riparian marshes with vegetation of reed or sedges where the host plant great water dock or related dock species are growing.
In the Netherlands found in the area of old peat cuttings on floating reed or sedge beds. This is the secondary vegetation that develops after peat cutting, so the species has switched from its original natural environment that disappeared about two centuries ago to the recent anthropogenic one.
In Luxembourg and Wallonia the large copper appeared in the 1940’s in the river valleys near the French border and has since then spread.
Butterfly is on the wing in the Netherlands from end-June until mid-August and peaks mid-July. In Belgium and Luxemburg from mid-May until early-September and peaks early-June and mid-August. The required heat sum is 700°d and the maximum tolerated 1800°d. In cool climates the species is restricted to sites with a warm micro-climate, in warm climate to sites with a cool and humid micro-climate of a tall vegetation.
The species is known from sub-maritime and continental climates, amplitude 7 to 18, corresponding climate windows are 26 and 38 weeks.
The larva feeds on dock species, not on sorrel and in the Netherlands nearly exclusively on Rumex hydrolapathum, in Belgium and Luxembourg on R. crispus and occasionally on R. obtisifolius.
The Dutch large copper was discovered near Wolvega in Friesland in 1915. The species occurs now only in the reserves Rottige meenthe, Weerribben and De Wieden; the small overwintering larvae survive well with the winter cutting of the reed beds.
About 1972 the species appeared spontaneously in the Makkumerwaard, a large reed bed on a sandbank in Lake IJsselmeer, 40 km from its nearest population. However, in spring of 1985 the whole area was destroyed during the thaw by drifting ice from IJsselmeer; the ice floes piled up to the top of the Frisian sea embankment.
Detailed studies on population dynamics were carried out in the Rottige Meenthe reserve, Friesland by 10 year counts of larvae in May and eggs in August along a 1455 m transect between 1960 and 1970 and one on the spatial distribution, in an area of 1500 ha in the Weerribben reserve, Overijssel in May 1968. The transect counts showed a high population level at the beginning but a sharp decline during 1962-1963, because of two bad summers in succession; the extinction seemed a realistic possibility.
However, the population recovered slowly up to 1970. The average larval mortality counted from egg to last instar larvae was about 98%. The distribution survey in the Weerribben showed that the larvae could be found in local concentrations but also that there were large areas with no larvae. This pattern was obviously correlated with the structure of the landscape. The average larval density was 0.16 per ha; however, in some places 0.34 and 0.46 per ha were recorded. These figures were typical for a low density species (Bink 1972).
The parasites observed during this survey were the egg parasite Trichogramma evanescens, a chalcidoid micro-wasp, and the ichneumonid wasp Hyposoter placides. The former needs egg-overwintering species to survive the winter, the latter attacks the second instar larvae in August and the mummified larvae will appear the following May.
Both parasites occur irregularly, there were years when none could be detected. The average incidence was about 2% but once a 20% incidence was observed. The wasp Hyposoter placides is a common parasite that occurs widely in Europe, from southern France to Estonia; it is a specialist attacking second instar Lycaeninae larvae and kills its host when the larva reaches about 15 mm in length.
Ecological characteristics
Behaviour over time
Overwintering: small larva third instar, 3-4 mm in size, hidden under the wilted leaf of the host in the litter-layer. The larva changes its colour from green into reddish brown when it is in diapause.
Reproduction: oviposition starts after 3-5 days when the body contains 150 (124-175) eggs. Observed production in breeding experiments 4.7 times as much.
Larval feeding periods: in late summer 33 (19-46) days, in spring 53 (43-66) days. In the case of a second brood 25 (18-40) days in summer.
Generations: one in the Netherlands and other countries at the same latitude, in the warmer southern parts as in Luxemburg and Wallonia two, sometime three.
Spreading of risk: extension of larval diapause of up to six weeks in the spring.
Life cycle: egg 7 (5-11) days; larva 43-49 weeks or 3-5 weeks in summer brood; pupa 18 (10-24) days.
Life span of adult: long, 3 to 5 weeks.
 
Photographs: Frits Bink ©.
Behaviour in space
From stay-at-home to migrant: stay-at-home, spatial requirement considerable; however, in the case of Luxemburg and Wallonia modest.
Finding a mate: male perches, the nomadic female seduces the male.
Orientation in the landscape: edge-like structures like river banks and sharp gradients between short and tall grass vegetation.
Oviposition: two to five eggs together on the upper or under side of a mature leaf.
Defence
Threats from other organisms: no defence, larva and adult very vulnerable to avian predation.
Myrmecophily: very little, ants inspect the larvae only for a short time.
Threats from the environment: vulnerable, larva and adult live in a humid and mild micro-climate, in the Benelux. The larva may survive inundation in winter and early spring.
Feeding habits
Adult: nectar of all kinds of flowers such as thistle, purple loosestrife and also honeydew produced by aphids living on reed.
Larva: in summer and autumn larvae feed on mature leaves, in spring they have a preference for young shoots.
Larval foodplants
Plant species: Polygonaceae, Rumex crispus, R. hydrolapathum, occasionally also R. obtusifolius, outside the Benelux R. aquaticus, R. sanguineus and others.
Journal
Breeding experiment based on specimen from Kalenberg, Netherlands:
29 July 1982: old female captured.
2 August: died after it laid 115 eggs.
8 August: eggs were hatching.
21 August: larvae in L3.
27 August: all larvae in diapause.
Overwintered outdoors.
3 April 1983: no activity observed.
17 April: about 10 larvae feeding.
20 April: 16 larvae active, three still in diapause.
3 May: last larva started to feed.
3 June: first pupa.
24 June: two adults appeared. Weight of the pupae: 375 ± 49 mg (n=10).
12 July: end of the oviposition, 7 females dissected which had produced on the average 714 eggs per female. See Table 33-1.
Exceptional second brood:
19 July 1983: 16 larvae of the offspring did not enter diapause thus starting a 2nd generation.
2 August: larvae of second brood pupating, the remaining small larvae entered diapause.
13 August: pupae weighed: “female” pupae 221 ± 24 mg (n= 7), “male” pupae 155 ± 18 mg (n=9).
17 August: first adults of the second brood appeared in greenhouse.
26 September: 2nd brood adults had mated. The butterflies produced later on 160 ± 72 in 15 egg-laying days, lifespan butterflies about 19 days.
16 October: all the remaining small larvae are in diapause and overwinter. The experiment on survival starts with these larvae, see Table 33-2.
Overwintered larvae:
7 March 1984: first larvae active in greenhouse.
8 March: larval survival examined (see Table 33-2).
19 March: larvae had started to feed.
16 April: some larvae in L4, some in diapause.
2 May: some larvae nearly full grown, smallest ones at end of third instar and changes its colour into green.
21 May: two pupae present.
12 June: adults appeared.
13 July: last pupa hatched, male.
Table 33-1. Results of dissections
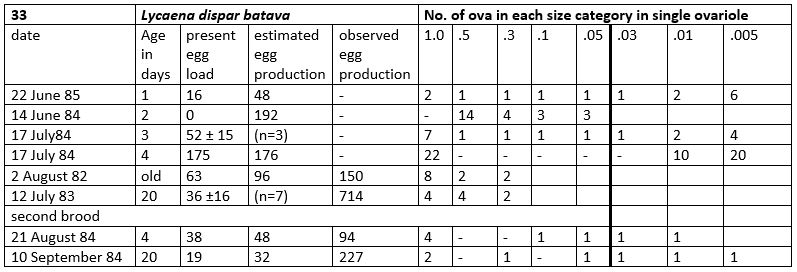
Table 33-2. Overwintering mortality of larvae, result from examination on 8 March 1984
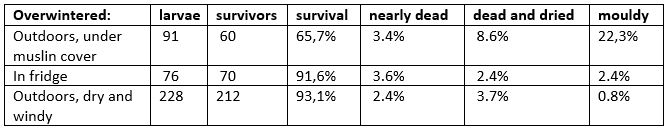
Table 33-3. Collection and observation localities
L. dispar carueli
B, Ethe, 240 m, 49° 36’ 25”N – 5° 35’ 50”E; 11 July 1984.
F, Lorraine, Jaulny 209 m, 48° 57’ 55”N – 5° 52’ 52”E; 8 July 2006.
L. dispar rutila
EST, Endla Nature Reserve, 58° 52 34”N – 26° 16’21”E; 10 July 1999.
EST, Kuresoo, 58° 28’ 00”N – 25° 11’ 59”E; 8 July 1999.
EST, Viruna 58° 28’ 51”N – 23° 58’ 32”E; 5 July 1999.
PL, Szczecin 53° 19’ 44”N – 14° 30’ 53”E; 7 July 1969, 14 July 1969.
L. dispar burdigalensis
F, Blanquefort 44° 55’ 02”N – 0° 36’ 11”W; 27 August 1964, 13 August 1968.
L. dispar batava
NL, Bancopolder 52° 52’ 49”N – 5° 41’ 05”E; 3 July 1966, 24 July 1969, 31 May 1981 (larvae).
NL, Rottige Meenthe, transect from 52° 50’ 12”N – 5° 54’ 01”E to 52° 49’ 43”N – 5° 54’ 01’E; 1960-1970.
NL, Weerribben, mapping, 52° 47’N – 5° 56’E; 15 April 1968 till 10 June 1968.
NL, Weerribben, Kalenberg 52° 46’ 45”N – 5° 56’ 16”E; 29 July 1982.
Fig. 33-1. Lycaena dispar batava phenogram adapted from Bos et al. 2006: 140 (dark part) & L. dispar carueli from Fichefet et al. 2008: 115 (lighter part).
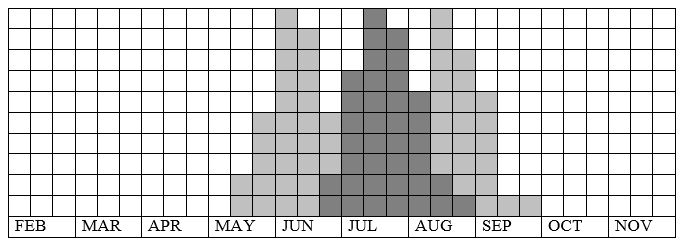
Fig. 33-2.Lycaena dispar, habitat characteristics.
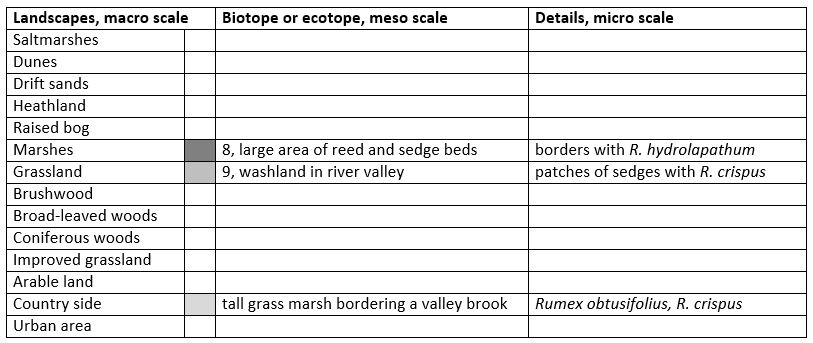
Fig. 33-3. Lycaena dispar, climate matrix, heat-sums 700 - 1800°d.
|